Markagunt Plateau Spruce Beetle Outbreak
During the early 1990s, endemic spruce beetle populations built to epidemic (outbreak) levels on the Markagunt Plateau in southern Utah. Although native to our western forests, the spruce beetle’s life cycle necessarily requires the death of the host tree, thus the meaning of their genus, Dendroctonus or tree killers! By the mid-2000s virtually all of the Engelmann spruce on the Markagunt Plateau had been killed (DeRose & Long 2007), radically changing the forested environment, and making this the most severe documented bark beetle outbreak in the state of Utah. In the wake of the spruce mortality caused by the outbreak, Ph.D. student Justin DeRose, advised by Dr. Jim Long, initiated a research project that aimed to elucidate the possible causes, patterns of spread, and history of disturbance, in order to better inform our understanding of Engelmann spruce disturbance ecology. The research also revealed important insights regarding the management of forests for their resistance and resilience to spruce beetle outbreaks.
Project Support

Spruce Beetle Outbreak Origin
Forest Stand Conditions
Just why did a spruce beetle outbreak develop and spread on the Markagunt Plateau? This outbreak was not the first recorded outbreak in the state. Outbreaks in the early 1900s on the Aquarius Plateau, and in the late-1980s have been reported in the literature. It is generally understood that spruce beetle outbreaks likely originate in areas of substantial windthrow, where beetle populations increase rapidly. They also reproduce well in dense stands of large, stressed spruce. Reconstructed structure and composition of live Engelmann spruce at the time of outbreak initiation revealed that the vast majority of Engelmann spruce stands on the Markagunt Plateau had incredibly high susceptibility to beetle attack—indicated by very dense stand conditions (high spruce basal areas) in combination with very large mean tree sizes (DeRose & Long 2012a). In addition, so many of the spruce stands were in such advanced stages of succession that commonly associated species that are not hosts to spruce beetle, such as subalpine fir and quaking aspen, were rare. The purity of spruce is a consequence of having not had been disturbed for many centuries (DeRose & Long 2012b).

Environmental Conditions
Drought is often implicated in driving bark beetle outbreaks through its direct effect on trees that become drought-stressed, which reduces vigor. However, during the early 1990s there were temperature anomalies in southern Utah (DeRose & Long 2012) that may have increased the likelihood for spruce beetle population growth (increasing growing season temperatures) and survival (ameliorated winter temperatures). An explicit assessment of the effect of drought vs. temperature on the spruce beetle outbreak revealed that temperature played a direct role in decreasing spruce vigor, likely making them more susceptible to beetle attack. When considered in combination with the direct control temperature has on the growth of beetle populations, and their presence in spruce forests across the west (DeRose et al. 2013), it seems likely that increasing temperatures played a role in the establishment and development of the Markagunt Plateau outbreak.
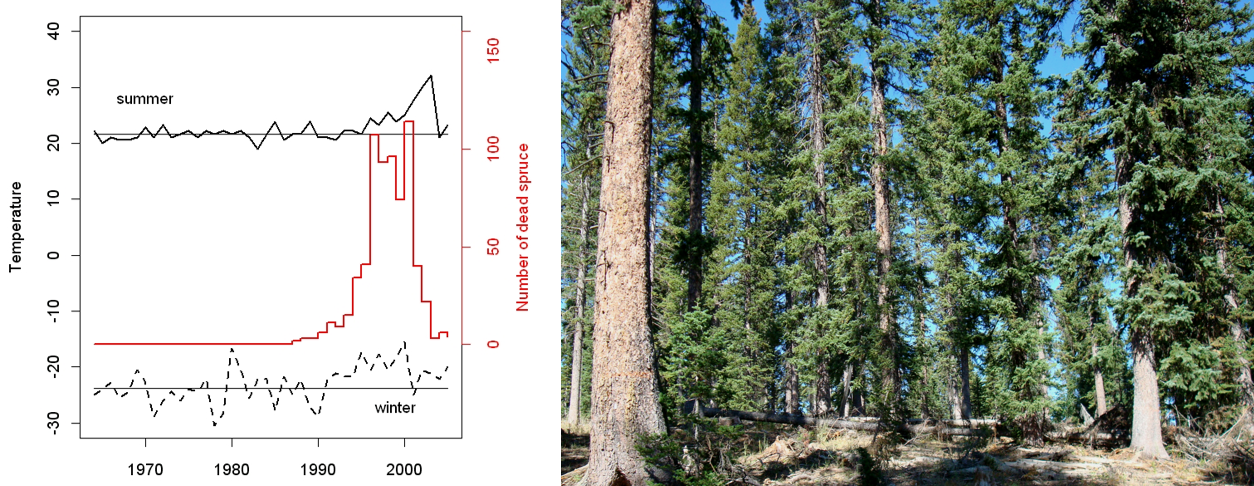
Outbreak Pattern
While no single starting point has been pinpointed for the Markagunt Plateau outbreak, the rise and spread of the outbreak was reconstructed using dendrochronological (tree-ring) techniques by dating the death of each tree across the plateau. These same techniques were also used to infer the exact timing of bark beetle attack on the very few, remaining live spruce (4.8%), providing a second line of evidence for our measurements of dates-of-death. Results from mapping dates-of-death across the plateau indicated that as the beetle populations increased, multiple, independent ‘hot spots’ of mortality emerged on the landscape—and over the course of many years these mortality pockets coalesced—until the host resource was exhausted (DeRose & Long 2012a, DeRose et al. 2011). The emergence of morality hot spots likely represents the ‘start’ of the outbreak, and these occurred predictably in areas of dense, pure, large spruce; mortality eventually spread to nearly all the available spruce on the landscape. Of the small percentage of spruce that did survive the outbreak, it appears they did so by rapidly creating resin ducts in their tree rings, which helped to deliver resin to beetle attack sites faster—allowing the trees to push the beetles out (DeRose et al. 2017).
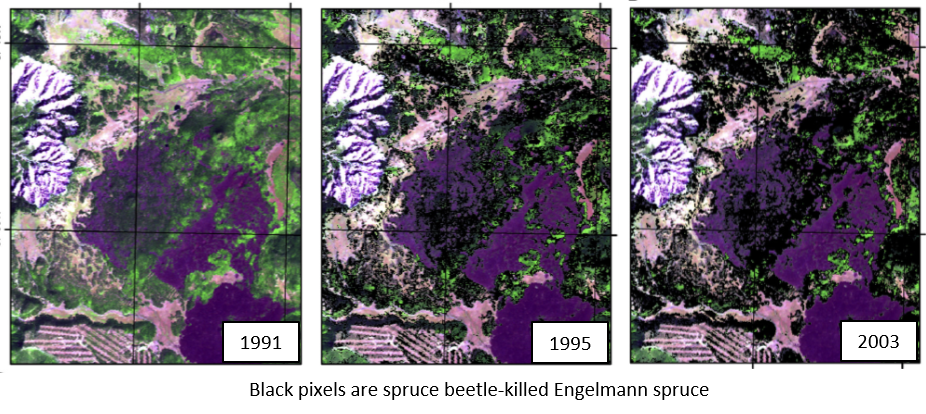
Disturbance History Reconstruction
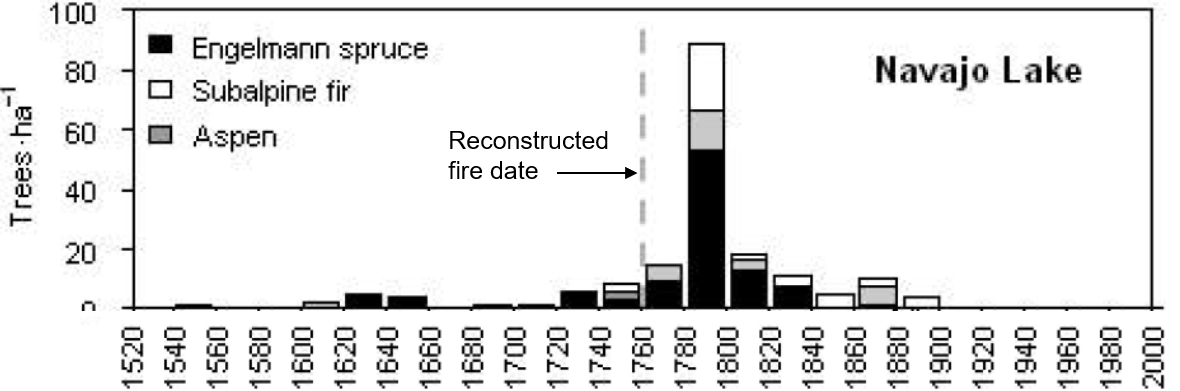
Are spruce beetle outbreaks of this magnitude and severity a normal part of Engelmann spruce disturbance regimes? Questions like these can be answered using dendrochronological techniques to infer the origins of the forests that eventually succumbed to the outbreak. Using these methods, the history of disturbance was reconstructed for dozens of forested stands across the outbreak area. Results revealed that all of the stands in question had originated from large, severe, stand-replacing fires, and not previous beetle outbreaks (DeRose & Long 2012b). This means that the jury is still out on whether beetle outbreaks of this size are within the natural range of variability.
Disturbance Interactions
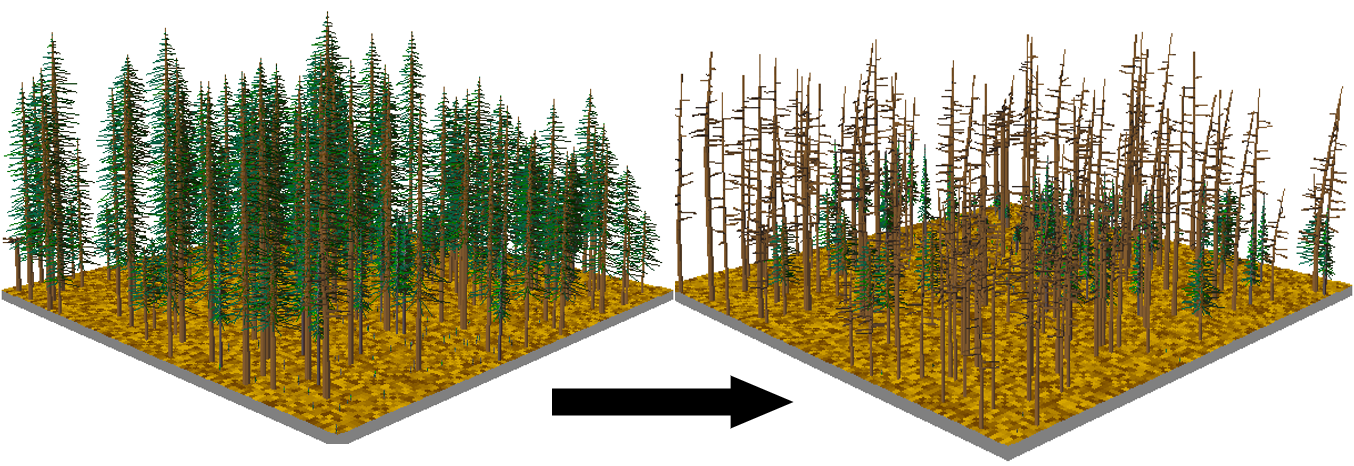
The sheer number of standing dead trees after the spruce beetle outbreak has the appearance of creating a worsened situation for subsequent fire risk. Indeed, one of the most commonly asked questions while conducting this research was ‘how soon do you think these forests will burn?’ While beetle outbreaks and wildfires can interact in space and time, where the occurrence of one can increase or decrease the occurrence of the other, the effect of the Markagunt Plateau outbreak on subsequent fire risk was counterintuitive. Post-outbreak, fire risk—estimated by calculating potential fire behavior—was actually lower after the outbreak (DeRose & Long 2009). How can this be? Fires are exceedingly rare in this high-elevation spruce ecosystem, re-occurring only every ~270-679 years over the last ~8,000 years (Morris et al. 2013). The likelihood of large, stand-replacing fires in cool, moist ecosystems like this spruce forest, is controlled by climate, and directly related to extreme drying typically associated with drought. Extreme drought lowers the moisture content of fuels, especially conifer needles, so low that they become incredibly flammable, should a fire start. The reduction of fine fuels, post-outbreak, led to lower potential fire behavior than conditions prior to the outbreak, reducing the possibility of stand-replacing crown fire (DeRose & Long 2009). It is interesting to note that, despite the fact that it has been approximately 351 years since the last stand-replacing fire on the Markagunt Plateau (Morris et al. 2013), the southern flank of the very large Brian Head Fire of 2017 did not make it into what were once pure, spruce-dominated forests prior to the spruce beetle outbreak.
Regeneration Ecology
One of the primary concerns after the Markagunt Plateau outbreak was the future of these forests—will there be regeneration failures, species shifts, forest type changes. Careful study revealed that the outbreak changed the drivers of Engelmann spruce regeneration, such that understory spruce that did not succumb to the bark beetle (advance regeneration) exhibited different strategies than spruce established after the outbreak (seedlings, Pettit et al. 2019). In addition, the presence of both types of regeneration strategies was highly variable across the landscape making management for them difficult. The amount of regeneration of all Engelmann spruce across the Markagunt Plateau was very low, while subalpine fir and aspen were much more prevalent, their numbers were highly variable (DeRose & Long 2010). The amount of regeneration for all species was somewhat predictable by their presence in the overstory, pre-outbreak (DeRose & Long 2010), which has been found to be common in spruce systems around the world (Zeppenfeld et al. 2015). In stands that were not pure spruce, subalpine fir and aspen are likely to dominate, and in stands where aspen was a larger component of the pre-outbreak composition, post-outbreak management activity is likely to successfully regenerate aspen (Britton et al. 2016).

Management Implications
Forest managers express concerns about the future of Engelmann spruce forests that have succumbed to spruce beetle outbreaks, and wonder if they can do anything to mitigate the effect of spruce beetles in forests yet affected by large-scale outbreaks. After an outbreak, foresters have the opportunity to salvage spruce, but more importantly to artificially regenerate by planting spruce. This stage in stand development is also the ideal time for them to consider augmenting their planting with non-host species, for example, Douglas-fir, white fir, limber pine, ponderosa pine, that might be more adapted to the site as climate changes. Increasing species diversity will decrease both the likelihood and severity of future spruce beetle outbreaks (DeRose & Long 2012a). In forests not yet effected by an outbreak, it is only a matter of time (Windmuller-Campione et al. 2021), and a prudent forester would not leave regeneration up to chance (Long et al. 2018). Fortunately, we have a better conceptual understanding of why and how beetle populations develop that can help us better prepare to for future outbreaks through the management of forest structure and composition. We can reduce susceptibility to spruce beetle via intermediate treatments like thinning, that reduces the risk of beetle populations transitioning from endemic to epidemic (outbreak) levels. However, these resistance approaches are short-term, and only buy time until the inevitable outbreak occurs (DeRose & Long 2014). Resilience approaches are recommended, as they focus on building age-class diversity of Engelmann spruce across the landscape, that is, multiple stands, so that when the outbreak occurs, there will be more area of spruce that is not susceptible to spruce beetle (Windmuller-Campione et al. 2015, 2021).

Literature
Britton, J.B., R.J. DeRose, K.E. Mock, and J.N. Long. 2016. Herbivory and advance reproduction influence quaking aspen regeneration response to management in southern Utah, USA. Canadian Journal of Forest Research 46(5):674–682. (link)
DeRose, R.J., M.F. Bekker, and J.N. Long. 2017. Traumatic resin ducts as indicators of bark beetle outbreaks. Canadian Journal of Forest Research 47:1168-1174. (link)
DeRose, R.J. and J.N. Long. 2014. Resistance and resilience: a conceptual model for silviculture. Forest Science 60:1205-1212. (link)
DeRose, R.J., B.J. Bentz, J.N. Long, and J.D. Shaw. 2013. Effect of increasing temperatures on the distribution of spruce beetle in Engelmann spruce forests of the Interior West, USA. Forest Ecology and Management. 308:198–206. (link)
DeRose, R.J., and J.N. Long. 2012a. Factors influencing the spatial and temporal dynamics of Engelmann spruce mortality during a spruce beetle outbreak. Forest Science. 58(1):1–14. (link)
DeRose, R.J. and J.N. Long. 2012b. Drought-driven disturbance history characterizes a southern Rocky Mountain subalpine forest. Canadian Journal of Forest Research 42:1649-1660. (link)
DeRose, R.J., J N. Long, and R.D. Ramsey. 2011. Combining dendrochronological data and the disturbance index to assess Engelmann spruce mortality caused by a spruce beetle outbreak in southern Utah, USA. Remote Sensing of Environment. 115(9):2342–2349. (link)
DeRose, R.J., and J.N. Long. 2010. Regeneration response and seedling bank dynamics of a Dendoctronus rufipennis-killed Picea engelmannii landscape. Journal of Vegetation Science. 21(2):377–387. (link)
DeRose, R.J. and J.N. Long. 2009. Wildfire and spruce beetle outbreak: simulation of interacting disturbances in the central Rocky Mountains. Ecoscience 16:28-38. (link)
DeRose, R.J., and J.N. Long. 2007. Disturbance, structure, and composition: spruce beetle and Engelmann spruce forests on the Markagunt Plateau, Utah. Forest Ecology and Management. 244(1–3):16–23. (link)
Long, J.N., M. Windmuller-Campione, and R.J. DeRose. 2018. Building resistance and resilience: regeneration should not be left to chance. Forests 9(5). (link)
Morris, J.L., A. Brunelle, R.J. DeRose, H. Seppä, M.J. Power, V. Carter, and R. Bares. 2013. Using fire regimes to delineate zones in a high-resolution lake sediment record from the western United States. Quaternary Research. 79(1):24–36. (link)
Pettit, J.M., S.L. Voelker, R.J. DeRose, and J.I. Burton. 2020. Spruce beetle outbreak was not driven by drought stress: evidence from a tree-ring iso-demographic approach indicate temperatures were more important. Global Change Biology 26(10): 5829-5843. (link)
Pettit, J.M., J I. Burton, R.J. DeRose, J.N. Long, and S.L. Voelker. 2019. Epidemic spruce beetle outbreak changes drivers of Engelmann spruce regeneration. Ecosphere. 10(11):e02912 (link)
Windmuller-Campione, M.A., R.J. DeRose, and J.N. Long. 2021. Landscape-scale drivers of resistance and resilience to bark beetles: A conceptual susceptibility model. Forests 12(6): 798. (link)
Windmuller-Campione, M.A., D.H. Page, and J.N. Long. 2017. Does the practice of silviculture build resilience to the spruce beetle? A case study of treated and untreated spruce-fir stands in northern Utah. Journal of Forestry. 115(6):559–567. (link)
Windmuller-Campione, A.M., and J.N. Long. 2015. If long-term resistance to a spruce beetle epidemic is futile, can silvicultural treatments increase resilience in spruce-fir forests in the Central Rocky Mountains? Forests. 6(4). (link)
Zeppenfeld, T., M. Svoboda, R.J. DeRose, M. Heurich, J. Müller, P. Čížková, M. Starý, R. Bače, and D.C. Donato. 2015. Response of mountain Picea abies forests to stand-replacing bark beetle outbreaks: neighbourhood effects lead to self-replacement. Journal of Applied Ecology. 52(5):1402–1411. (link)
